 Нервная система состоит из двух типов клеток — нервных (нейроны) и глиальных (нейроглия). К нейроглии относятся астроциты, микроглия, миелинобразующие клетки (олигодендроциты ЦНС и шванновские [1] клетки периферических нервов).
Нервная система состоит из двух типов клеток — нервных (нейроны) и глиальных (нейроглия). К нейроглии относятся астроциты, микроглия, миелинобразующие клетки (олигодендроциты ЦНС и шванновские [1] клетки периферических нервов).
По отношению к нейронам глиоциты выполняют трофическую, опорную и изолирующую (электрический изолятор) функции.
Нейрон — основная структурно-функциональная единица нервной системы. Число нейронов в мозге человека превышает 100?109. Основная функция нейронов — генерация, передача и интеграция нервных импульсов.
В нервной клетке различают тело (перикарион) и отростки — дендриты и аксон (рис. 5–1). Как правило, нейрон имеет несколько дендритов и один аксон. Отростки нейрона и поверхность перикариона участвуют в образовании синапсов. Нейроны — возбудимые клетки — осуществляют передачу электрических сигналов в пределах самой нервной клетки (а между нейронами при помощи нейромедиаторов в синапсах) и этим обеспечивают способность мозга к переработке информации.
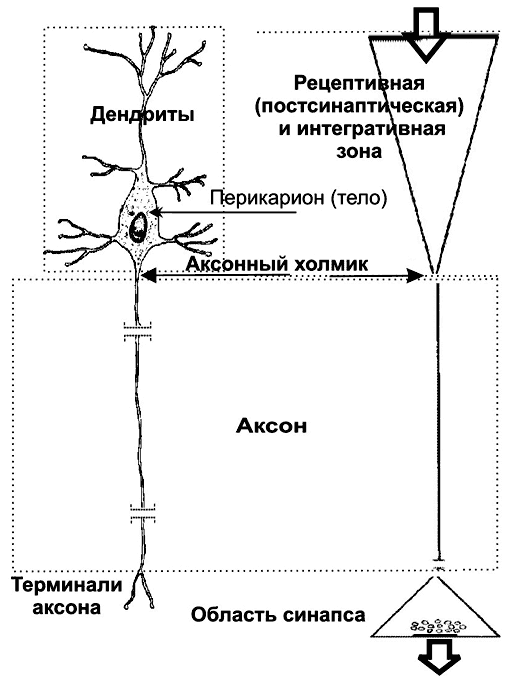
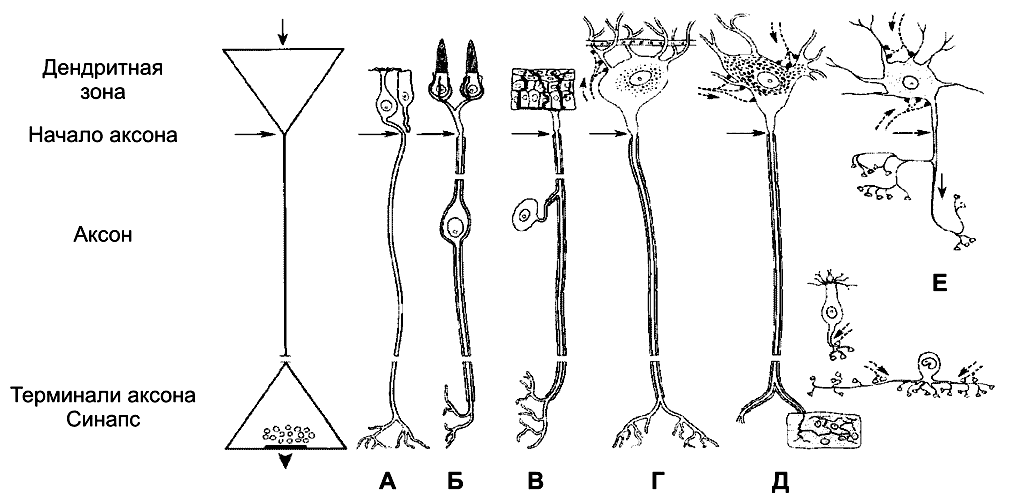
Рис. 5–1. Многоотростчатый нейрон [11]. Тело клетки (перикарион) содержит ядро. От перикариона отходят отростки. Один из них — аксон, все другие — дендриты. Справа и сверху вниз: функциональные зоны нервной клетки — рецептивная (дендритная), аксон (область проведения ПД), концевых разветвлений аксона (пресинаптическая).
Дендриты — ветвящиеся отростки, заканчивающиеся вблизи от тела нейрона. Дендриты имеют многочисленные шипики, значительно увеличивающие их поверхность. В функциональном отношении дендриты — рецепторная зона (или постсинаптическая область) регистрации сигналов от других нервных клеток. Поверхностная мембрана дендритов и перикариона одного нейрона головного мозга образует до 20 000 синаптических контактов с другими нейронами. Другими словами, в поверхностную мембрану, покрывающую дендриты и перикарион, встроено множество рецепторов к нейромедиаторам, секретируемым другими нервными клетками.
Тело нейрона содержит ядро, комплекс Гольджи, [2] гранулярную эндоплазматическую сеть, митохондрии, лизосомы, элементы цитоскелета. Синтез белка происходит только в перикарионе.
Интегративная зона — тело нейрона вместе с дендритами и областью отхождения аксона, в этой зоне интегрируются локальные изменения МП, происходящие в местах синаптических контактов.
Аксонный холмик (начальный сегмент аксона) — место, где начинается аксон и происходит генерация ПД.
Аксон — длинный отросток, как правило, не ветвящийся. Объём аксона может достигать 99% суммарного объёма нейрона. Длина аксона может быть значительной — десятки сантиметров. Основной функцией аксона является проведение ПД.
Аксонный транспорт. Поскольку в аксоне и нервных окончаниях практически нет рибосом, необходимые для деятельности всей нервной клетки белки синтезируются в перикарионе, а затем транспортируются по аксону посредством аксонного транспорта.
Терминали аксона (концевые разветвления) принимают участие в образовании синапсов с другими нейронами, мышечными и секреторными клетками. Терминали заканчивается утолщениями, содержащими синаптические пузырьки (везикулы).
Классификации нейронов
Существует множество классификаций нервных клеток, так как нейроны отличаются по размерам и форме перикариона, числу отростков, их синаптическим связям, характеру ветвления дендритов, электрофизиологическим характеристикам, химии нейромедиаторов, позиции в нейронных сетях и множеству других характеристик. В зависимости от класса выполняемой функции выделяют афферентные (чувствительные, сенсорные), эфферентные (двигательные, моторные) и вставочные нервные клетки (ассоциативные нейроны, или интернейроны).
Афферентные нейроны проводят возбуждение от рецепторов периферических органов в структуры ЦНС.
Эфферентные нейроны осуществляют передачу сигналов от ЦНС к органам-эффекторам (мышцам и железам).
Ассоциативные нейроны проводят возбуждение между нейронами.
Морфологическая классификация. По количеству отростков нейроны разделяют на псевдоуниполярные, биполярные и мультиполярные.
Псевдоуниполярные нейроны имеют один короткий отросток, который разделяется на некотором расстоянии от сомы на два длинных — дендрит и аксон. К псевдоуниполярным относятся нейроны сенсорных ганглиев спинного мозга.
Биполярные нейроны имеют один дендрит и один аксон. Этот вид нейронов встречается в периферическом отделе зрительного, обонятельного и слухового анализаторов.
Мультиполярные нейроны имеют один аксон и несколько дендритов, это наиболее распространённый вид нейронов. К ним относятся мотонейроны спинного мозга.
Исходя из характера электрогенеза в различных зонах нервных клеток (рецептивная и интегративная зоны, начальный сегмент аксона, аксон, терминали аксона), предложена универсальная классификация (рис. 5–2, см. также рис. 5–1). Эта классификация учитывает направление (вход и выход) сигналов в пределах одной нервной клетки, но находящейся в составе нейронных сетей или цепочек, а также характер мембранного электрогенеза (изменения МП в рецептивной и пресинаптической зонах и передача ПД по аксону).
Рис. 5–2. Универсальная классификация частей различных типов нейронов [11]. Дендриты — рецепторная зона — возбуждающий или тормозящий вход. Перикарион с дендритами — интегративная зона. Аксон — проводящая возбуждение (ПД) часть нейрона, его концевые разветвления образуют синапсы (выход) с соседними клетками. А–Г — афферентные (чувствительные) нейроны; Д — мотонейрон; Е — интернейроны.
Мембранный потенциал покоя
Мембранный потенциал (МП) покоя — разность электрических потенциалов между внутренней и наружной поверхностью мембраны в состоянии покоя (при отсутствии раздражителей) — рассмотрен в главе 2 (раздел «Мембранный электрогенез»). МП покоя нервной клетки составляет –70 мВ, и его величина достаточна стабильна. Изменения величины МП (рис. 5–3) описывают специальными терминами:
- деполяризация — уменьшение величины МП;
- реполяризация — возвращение величины МП к исходному уровню;
- гиперполяризация — увеличение МП по сравнению с исходным уровнем.
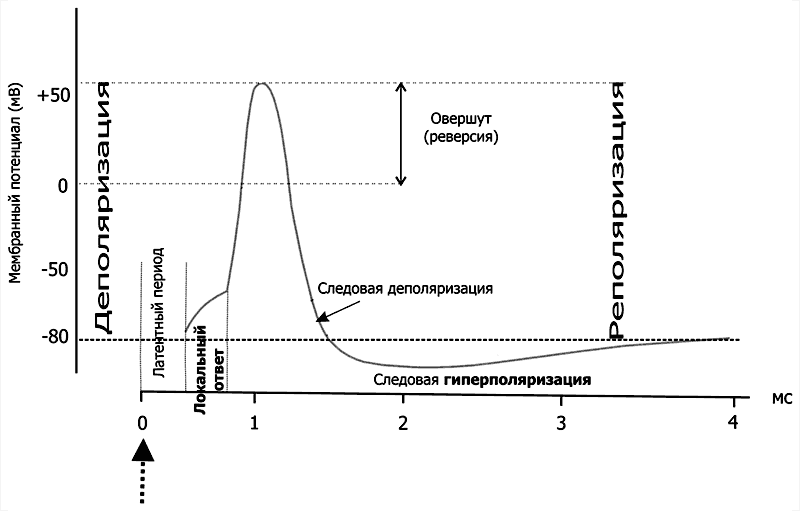
Рис. 5–3. Изменения мембранного потенциала и потенциал действия. Вертикальная стрелка в нижней части рисунка — момент появления раздражающего стимула, на отметке –80 мВ — исходный уровень МП.
Все изменения МП в дендритной зоне нервной клетки известны как локальные ответы, затухающие на протяжении нескольких микрометров от места возникновения. В этой зоне постоянно возникают десятки и сотни локальных ответов (постсинаптические потенциалы), суммирование которых в виде изменений МП достигает области начала аксона (аксонного холмика). Помимо локальных ответов, возбудимые мембраны нервных клеток в области аксона могут генерировать потенциалы действия (ПД, спайки, нервные импульсы, рис. 5–3 и 5–4). Существенное значение при генерации ПД имеет представление о пороге раздражения.
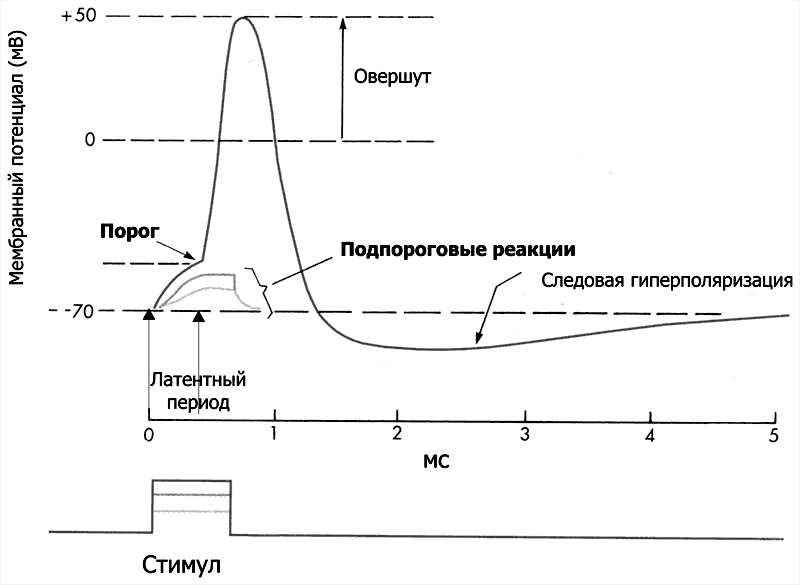
Рис. 5–4. Мембранный потенциал и потенциал действия. На отметке –70 мВ — исходный уровень МП; значение порога показано стрелкой в левой части рисунка, приведены записи подпороговых изменений МП.
Локальные ответы, порог раздражения и потенциал действия
Нервная клетка реагирует изменениями МП в ответ на раздражающие (в том числе электрические) стимулы либо локальными (подпороговыми) реакциями, либо генерирует ПД.
Локальные ответы. Стимулы, величина которых меньше определённого порога, вызывают локальные ответы, затухающие вблизи места раздражения. Эти локальные ответы могут быть как деполяризующие (возбуждающие) мембрану, так и гиперполяризующие (тормозные).
Потенциал действия. Деполяризующие стимулы, достигшие порогового значения, вызывают развитие возбуждения в виде ПД. ПД распространяется по мембране нейрона без затухания (без декремента).
Законы раздражения нервной клетки
Генерация ПД подчиняется законам «всё или ничего», «сила–время» и аккомодации.
Закон «всё или ничего»
При достижении пороговой силы раздражающего стимула дальнейшее увеличение его интенсивности или продолжительности раздражения не изменяет характеристик ПД. Такая форма реакции нервной клетки на раздражение получила название «всё или ничего» (либо ПД возникает, либо нет).
Закон силы–времени
Эффект раздражителя на нервную клетку зависит не только от силы раздражителя, но и от времени, в течение которого он действует; так, чем больше сила тока, тем меньше времени он должен действовать, чтобы возник процесс возбуждения, и наоборот. Соотношение силы и длительности действия раздражителя может быть выражено в виде гиперболической кривой (рис. 5–5).
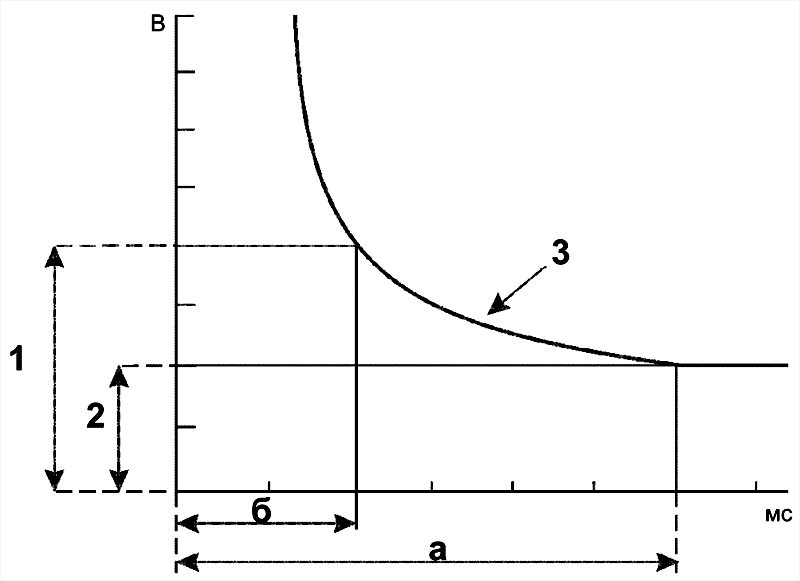
Рис. 5–5. Кривая «силы–времени» [21]. 1 — реобаза (минимальная сила раздражителя, способная вызвать ПД); 2 — удвоенная реобаза; 3 — кривая «силы-времени»; а — полезное время (минимальное время, в течение которого ток, равный одной реобазе, вызывает генерацию ПД); б — хронаксия (минимальное время, в течение которого ток, равный двум реобазам, вызывает ПД).
Из графика на рисунке 5–4 видно, что усиление раздражителя приводит к уменьшению минимального времени раздражения, но не бесконечно. При очень малых значениях длительности стимула кривая «силы-времени» становится почти параллельной к оси ординат. Точно также невозможно вызвать генерацию ПД длительным, но очень малым по амплитуде раздражающим стимулом.
Закон аккомодации
Раздражающее действие тока (рис. 5–6) зависит не только от амплитуды электрического сигнала, но и от скорости (крутизны) его нарастания во времени: чем быстрее это нарастание, тем сильнее выражено раздражающее действие тока. При действии медленно нарастающей силы раздражителя генерации ПД не происходит, так как возбудимая клетка «приспосабливается» к действию этого раздражителя, что и получило название аккомодации.
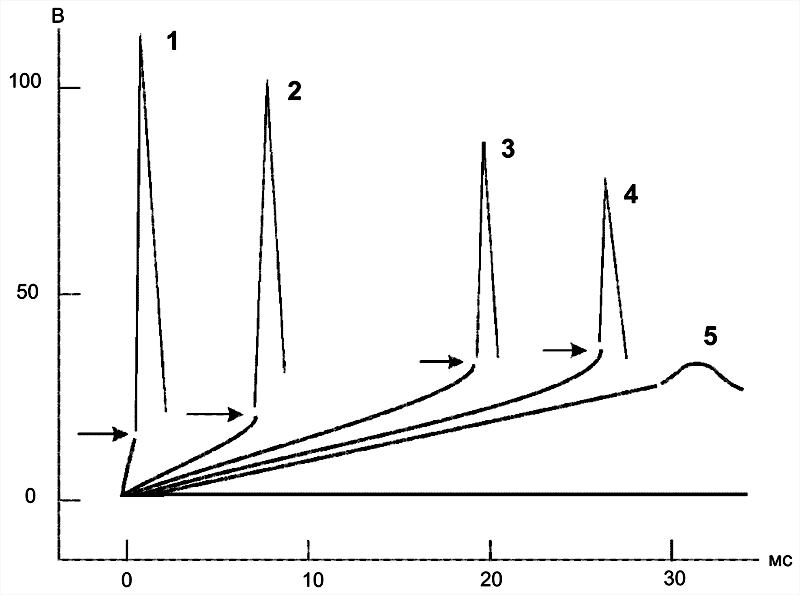
Рис. 5–6. Аккомодация мембраны нервной клетки к действию токов различной крутизны нарастания [21]. На графике показано изменение критического уровня деполяризации и амплитуды ПД (1-5) при раздражении нервной клетки линейно нарастающими токами различной крутизны.
Орлов Р.С., Ноздрачёв А.Д. Нормальная физиология : Учебник. - М.: ГЭОТАР-Медиа, 2009. - 688 с. - Нейроны / Глава 5. Физиология нейронов. С. 69-73.
[1] Шванн Теодор (Schwann T.), немецкий гистолог и физиолог (1810–1882); вместе с М. Шульце создал клеточную теорию (1839 г.); в 1836 г. открыл пепсин, в 1838 г. опубликовал первую работу по строению миелиновой оболочки.
[2] Гольджи Камилло (Golgi C.), итальянский гистолог (1843–1926). Разработал метод окрашивания ткани, основанный на импрегнации соединениями металлов. Метод был развит Рамон-и-Кахалем и его учениками, что позволило описать строение практически всех отделов нервной системы. За разработку этого метода Гольджи в 1906 г. был удостоен Нобелевской премии по медицине вместе с Сантъяго Рамон-и-Кахалем.