 Старая истина, что все объективные процессы протекают во времени и пространстве, на каждом этапе развития науки дополняется новым содержанием.
Старая истина, что все объективные процессы протекают во времени и пространстве, на каждом этапе развития науки дополняется новым содержанием.
В биологическую науку сегодня прочно проникает мысль, которую кратко можно выразить словами: «Ритм правит жизнью». Ритм можно увидеть в сокращениях сердца, в дыхании, в смене периодов двигательной деятельности организма, в сезонных гормональных перестройках, во влиянии 11-летнего цикла солнечной активности на биосферу... Существует многочисленная литература по биоритмам, и ее объем непрерывно возрастает.
Ритмы ЭЭГ
Но, пожалуй, наиболее наглядно роль ритмических процессов выступает при изучении биопотенциалов головного мозга. Уже первые записи электроэнцефалограмм (ЭЭГ), осуществленные в 20-х годах нашего столетия, показали преобладание в коре больших полушарий у человека ритма биопотенциалов с частотой 8—12 гц (Бергер, 1928). Он получил название альфа-ритм. Затем и другие буквы греческого алфавита стали использоваться для обозначения разных ритмов ЭЭГ. Так, слева от альфа расположился тета-ритм (4—7 гц), а еще левее — дельта-ритм (0,5—3 гц). Справа от альфа-ритма находится бета-ритм (13—20 гц), а еще правее — гамма-ритм (21 — 50 гц).
Не затрагивая сверхмедленных и слишком быстрых биопотенциалов мозга, можно считать, что главные частоты ЭЭГ располагаются в пределах от 0,5 до 50 гц.
Использование букв для обозначения ритмов биопотенциалов мозга не только удобно, но в определенной мере и знаменательно, ибо речь может идти о своеобразном «языке» мозговой деятельности, который удалось подслушать с помощью электронных приборов. Здесь работа, электрофизиолога похожа на исследования лингвиста-археолога, который открыл язык прошлой цивилизации. После открытия начинается самое сложное — расшифровка смысла языка, т. е. перевод на другой, более понятный язык.
Вырисовывалась гипотеза, что более медленные ритмы ЭЭГ связаны с процессом торможения в головном мозгу, а более быстрые — с процессом возбуждения. Однако многочисленные проверки этой гипотезы показали, что связь между основными нервными процессми и ЭЭГ сложнее и что только по данным ЭЭГ трудно судить о возбуждении или торможении. Требовались другие дополнительные методы исследования мозга, чтобы ответить на вопрос о преобладании в центральной нервной системе (ЦНС) тормозного или возбудительного процесса.
Попытки обнаружить соответствие между изменениями ЭЭГ и условно-рефлекторной деятельностью головного мозга хотя и показали определенную связь между этими явлениями, но прочной взаимозависимости не обнаружили.
Почти 30 лет назад одним из нас совместно с К. Л. Поляковым (М. Н. Ливанов, К. Л. Поляков, 1945) было выяснено, что выработка оборонительного условного рефлекса у кроликов сопровождается глубокой перестройкой фоновых биопотенциалов коры головного мозга. Бросались в глаза два обстоятельства: во-первых, происходит обширная (в пространственном отношении) перестройка ритмики ЭЭГ и, во-вторых, начинается она раньше появления двигательных компонентов условного рефлекса. Однако эти изменения ЭЭГ ослабевали и почти исчезали при укреплении временной связи. Как показали в дальнейшем многочисленные данные советских и зарубежных исследователей, диффузная (разлитая, охватывающая различные участки) перестройка биопотенциалов головного мозга при становлении условного рефлекса четко соответствовала фазе его генерализации (обобщенного ответа на разные сигналы), но и это мало помогало выяснению сущности явления.
Дистантная синхронизация биопотенциалов
Итак, попытки перевести «язык» биопотенциалов головного мозга на другие «языки» его деятельности хотя и вселяли некоторую надежду, но окончательного успеха не давали. Но, может быть, принцип чтения знаков ЭЭГ должен быть другим? Как говорит пословица, один в поле не воин. До сих пор чаще прислушивались к сольному «пению» отдельных участков мозга, а музыка, возможно, целиком раскрывается в хоровом многоголосье.
Возникло предположение, что в основе обширной диффузной перестройки корковых биопотенциалов лежит процесс пространственной соритмии или, как мы его чаще называем, процесс дистантной синхронизации колебаний биопотенциалов.
Его можно сравнить с четкой поступью парадных колонн, шагающих под звуки оркестра по широким просторам площади. Тогда фоновая многоритмичная ЭЭГ будет подобна толпе, идущей не в ногу. Такое предположение с необходимостью требовало включения в анализ биопотенциалов мозга не только их временных, но и пространственных параметров.
Изучение пространственной синхронизации биопотенциалов головного мозга потребовало создания новых приборов. Ранее описанные результаты были получены при регистрации ЭЭГ от нескольких (4, 8 иногда 16) пунктов головного мозга. В нашей лаборатории, используя новый метод регистрации ЭЭГ — электроэнцефалоскопию (М. Н. Ливанов, В. М. Ананьев, 1960), удалось зарегистрировать электрическую активность сразу от 50 до 100 точек коры головного мозга. Сопоставляя соритмичность электрической активности разных участков мозга, иными словами, определяя дистантную синхронизацию биопотенциалов, оцениваемую математически по величине кросскорреляционных коэффициентов (то есть по степени соответствия биопотенциалов в разных участках мозга), исследователи смогли более детально описать работу мозга.
Оказалось, что в головном мозгу человека, когда он находится в спокойном состоянии, относительно мало (около 2,8%) участков работают синхронно.
Нужно заметить, что речь идет о высоких значениях коэффициентов корреляции. Они могут изменяться от –1 до +1, а синхронно работающими считались участки мозга, для которых этот показатель равнялся 0,8—1,0. Длительность записи ЭЭГ, по которой определялись синхронность, или «эпоха анализа», равнялась всего 1,5 сек.
Через 15—25 сек после начала выполнения умственной работы (перемножение двух двухзначных чисел) число точек головного мозга, работающих синхронно, т. е. взаимосвязанно, увеличивалось в несколько раз.
Интересно, что такие точки располагались преимущественно в лобных долях, принимающих активное участие в интеллектуальных действиях человека. Кроме того, этот процесс более отчетливо проявлялся в левом полушарии, которое у большинства людей (правшей) доминирует над правым полушарием. Чем труднее решалась предложенная задача, тем обширнее и длительнее выявлялись эти изменения.
Если испытуемый принимал терапевтическую дозу аминазина (25 мг), который блокирует влияние глубинных отделов головного мозга (ретикулярной формации) на кору, то выполнение умственного задания замедлялось и соответственно уменьшалась синхронность в лобовых отделах коры.
Сотрудником нашей лаборатории Н. А. Гавриловой было обнаружено, что при психических заболеваниях (некоторые формы шизофрении, эпилепсии) даже в покое резко увеличивается число синхронно работающих точек мозга (особенно при выраженном систематизированном бреде), но их расположение сдвигается к срединным отделам полушария.
Во время умственной нагрузки число синхронно работающих точек мозга у таких больных иногда даже уменьшалось. Эти результаты показывают, что при психических заболеваниях значительно нарушается распределение нервных процессов в ЦНС.
Следовательно, запись и анализ электрической активности нескольких участков головного мозга показали, что таким образом можно получить дополнительную информацию о деятельности мозга. Вставал вопрос о физиологической значимости синхронности процессов, протекающих в различных пунктах головного мозга. Для чего эти пункты начинали работать в унисон?
Можно предполагать, что в дистантной синхронности колебаний биопотенциалов отражаются условия, которые необходимы для осуществления связи между соответствующими участками мозга. Чтобы детально проверить это предположение, понадобились исследования на лабораторных животных
Преимущества управляемого эксперимента
Временное отступление от непосредственного изучения работы головного мозга человека было на самом деле шагом вперед, так как новый методологический принцип потребовал экспериментальной расшифровки ряда деталей. Переход от электроэнцефалографии к электроэнцефалоскопии нельзя воспринимать как простое количественное увеличение числа точек мозга, с которых регистрируется ЭЭГ. Даже полуторасекундная запись биопотенциалов с 50 каналов потребовала бы месяца работы при ручном методе обработки результатов. Качественное изменение характера электрофизиологического эксперимента могло обеспечить только использование электронно-вычислительных машин (ЭВМ).
Являясь символом и орудием научно-технического прогресса, ЭВМ не только расширяют возможности обработки физиологических данных, но влияют и на форму эксперимента и на задачи, формулируемые экспериментатором.
В нашей лаборатории В. А. Кравченко был создан комплекс аппаратуры, куда входили 24 усилителя биопотенциалов, ЭВМ марки «Днепр» и аналого-дискретный преобразователь, позволяющий вводить ЭЭГ кролика в ЭВМ. ЭВМ в реальном времени при частоте опроса 100 гц по каждому каналу ЭЭГ подсчитывала синхронность биопотенциалов 20 точек головного мозга. Кроме данных ЭЭГ, в машину вводились и электромиграфические (ЭМГ) показатели, характеризующие деятельность мышц конечностей животного.
Здесь уместно вернуться к сравнению нейрофизиологии с лингвистикой. Поскольку движения уже давно и правильно считают отражением мозговой деятельности, выражение «мысли композитора были ярко выражены языком танца» имеет под собой научную основу. Правда, в описываемых опытах речь идет не о балерине, а о кролике, но и у него «двигательный язьщ» мозга является наиболее понятным и изученным. Вот почему «язык» движений сопоставлялся с «языком» биопотенциалов головного мозга.
Изучали условия перехода возбуждения со зрительного анализатора на двигательный, т. е. определяли изменения пространственной синхронизации ЭЭГ, предшествующие возникновению мышечного сокращения лапы кролика, при световом раздражении.
Широкие возможности открывало наличие на выходе ЭВМ устройства обратной связи с объектом. Это позволило осуществить принципиально новую форму исследования — управляемый эксперимент
Суть этого метода заключается в том, что ЭЭГ кролика, регистрируемая непрерывно в нескольких точках мозга, тут же непрерывно обрабатывается ЭВМ. По заданной программе, когда соотношения показателей ЭЭГ отвечают определенным условиям, сама ЭВМ (а не экспериментатор!) подает раздражение (в данном случае свет) и оценивает возникающую мышечную реакцию.
Таким образом, исследователь исключается из черновой экспериментальной работы и его функция заключается в планировании эксперимента, в составлении заданий ЭВМ, которая выполняет программы быстрее и лучше десятка лаборантов. Этот пример показывает не только новые возможности, открываемые научно-техническим прогрессом перед исследователями работы мозга, но и путь повышения производительности труда самих исследователей.
Однако главные преимущества управляемого эксперимента заключаются в возможности мгновенного вмешательства, так сказать, по ходу дела, в деятельность мозга на основа-нии принятой от него и обработанной ЭВМ обширной информации. Таким быстродействием сам человек не обладает.
Условия взаимодействия нервных центров
Но вернемся к конкретному эксперименту. Предварительно у скальпированного кролика с помощью прямого электрического раздражения отыскивали в двигательной области коры представительство той или иной конечности (слабое электрическое раздражение двигательного центра вызывает небольшое сокращение мышц лапы). Отведение биопотенциалов осуществляли от найденного двигательного центра, от 19 точек задней части этого же полушария, где располагаются корковые зрительные центры, и от мышц избранной ранее лапы.
В результате описанных экспериментов было точно установлено, что важным условием перехода возбуждения со зрительных центров на двигательные является высокая синхронность биопотенциалов в исследуемых точках коры головного мозга.
Эту синхронность, а вернее отражающие ее кросскорреляционные коэффициенты между двигательным и зрительным центрами коры, непрерывно вычисляла ЭВМ. Когда такой коэффициент, т. е. пространственная синхронизация процессов, достигал большой величины (0,8—1,0), то ЭВМ (с запаздыванием меньше 10 мсек) включала перед кроликом вспышку света. Если осуществлялось движение, то оно обязательно регистрировалось ЭВМ. Теперь уже можно было прямо решать, имеется ли связь между высокой синхронностью биопотенциалов исследуемых точек мозга и переходом возбуждения между ними.
Статистическая обработка многочисленных опытов показала, что при высокой синхронности биопотенциалов (от 0,8 до 1,0) чаще осуществляется движение на свет, чем при низких (от –0,2 до 0,2). Следовательно, синхронность протекания электрических процессов в двух отдаленных участках коры головного мозга действительно является одним из условий их взаимодействия.
Другое условие выявилось при анализе числа точек зрительной области коры, биопотенциалы которых были синхронны с биопотенциалами двигательного центра. Оказалось, что чем больше было таких точек, тем легче возбуждение переходило со зрительного анализатора на двигательный.
Все равно как, чем сильнее и чем больше людей аукается в лесу, тем больше вероятности, что за блуждавшийся откликнется. Однако дальнейшие исследования показали, что важна не только количественная, но и качественная сторона взаимодействия нервных центров.
Частотный анализ ЭЭГ
Частота и амплитуда — вот основные параменты кривых, которые видит экспериментатор на записях ЭЭГ. Правда, речь может идти и о форме каждой волны, о характере ее восходящей и нисходящей части, но к этим деталям вернемся позднее.
Итак, частота колебания ЭЭГ. Является ли она существенным признаком, или синхронность, соритмия любых колебаний обеспечивает переход возбуждения между регистрируемыми пунктами мозга независимо от частоты, на которой она возникла?
Сотрудник нашей лаборатории В. Д. Труш провел обширные исследования по частотному анализу ЭЭГ с использованием ЭВМ. В 150 отрезках ЭЭГ, в которых наблюдалась высокая синхронность биопотенциалов, 90% составляли отрезки, где преобладали ритмы 4–7 гц, т. е. тета-диапазона. В тех записях ЭЭГ, где синхронность была мала, доминирование тета-ритма встречалось только в 6% случаев.

Контуры мозга здорового испытуемого с электродами, с помощью которых регистрируются биопотенциалы мозга. Белые кружочки обозначают участки мозга, которые не обнаружили высокой корреляции (синхронности) с другими отведениями, кружочки с черточкой указывают на синхронные участки в лобной области, с крестиком — в задних отделах коры, а с точкой — в двигательной области коры. На первом контуре мозга (верхний слева) обозначены результаты, полученные в состоянии покоя испытуемого, на втором (верхний справа) — через 25 сек после предъявления арифметической задачи. Наконец, на четвертом контуре видна синхронность различных точек коры после решения задачи
Регистрация биопотенциалов головного мозга больного шизофренией при выраженном бреде преследования. Кружочки обозначают то же, что на предыдущем рис. Слева — состояние биопотенциалов мозга до предъявления арифметической задачи (покоем это состояние едва ли можно назвать). Процесс решения задачи (см. справа) не вызвал резких изменений биопотенциалов, как наблюдалось у здорового испытуемого
Кроме того, выяснилось, что для передачи возбуждения в коре решающее значение имеет не только наличие сходных ритмов в тета-диапазоне, но и строгое соответствие друг другу отдельных колебаний этих ритмов в определенных пунктах коры. Это соответствие отражается так называемой «когерентностью» (совпадением по частоте) и соотношением фаз колебаний. При низкой когерентности переход возбуждения затруднен.
Таким образом, синхронность биопотенциалов, площадь коры, ею охватываемая, выраженность колебаний в диапазоне тета-ритма и когерентность — вот те необходимые условия, которые определяют распространение, облегчают переход возбуждения в коре головного мозга.
Лингвист назвал бы эти условия правилами, с помощью которых можно осуществлять перевод с одного языка на другой. Причем правила эти используются пока только для перевода нескольких фраз.
Искусственное усиление тета-ритма
Если все обстоит именно так, то правомерен вопрос: а нельзя ли искусственно влиять на переход возбуждения, изменяя доминирующую частоту ЭЭГ? В опытах Ю. А. Холодова и А. В. Кориневского кроликам давали световые вспышки с частотой 2; 5 или 9 гц и регистрировали частоту движений задней лапы.
Напомним, что к тета-ритму из примененных частот относится только частота 5 гц. Оказалось, что при световых раздражениях с частотой в 5 гц движения возникали в 2 раза чаще, чем при других используемых частотах. Следовательно, искусственное усиление тета-ритма увеличивает число движений.
Такое усиление тета-ритма можно вызвать и прямым слабым электрическим раздражением коры. И в этом случае движения на свет возникали чаще. С другой стороны, когда на голову кролика влияли электромагнитным полем радиочастотного диапазона или постоянным магнитным полем, движения на свет возникали реже, так как в ЭЭГ кролика при действии вышеуказанных факторов преобладали медленные волны дельта диапазона.
Большой интерес вызывает гипотеза о возможности «навязывания» ритмов ЭЭГ с помощью соответственно модулированных электромагнитных полей. Иногда указывают на поразительное соответствие частоты альфа-ритма ЭЭГ человека определенной частоте колебаний геомагнитного поля.
Часто при этом ссылаются на мнение известного кибернетика Н. Винера, который предполагал, что биологическая автоколебательная система, как и техническая, «захватывает» частоту действующих на нее внешних периодических процессов.
В опытах было обнаружено, что мозг (речь идет о мозге кролика) с трудом покоряется частоте подаваемых раздражений. Электромагнитные поля, как уже говорилось, независимо от их частотной модуляции вызывают в ЭЭГ преимущественно дельта-ритм. Даже такой высокоразвитый анализатор, как зрительный, часто следует за ритмом вспышек только в диапазоне частот, соответствующих тета-ритму.
А если частота вспышек будет выше или ниже указанного диапазона, в ЭЭГ зрительной области коры больших полушарий усиливаются и задаваемые частоты и тета-ритм. В двигательной области коры при разной частоте светового раздражения всегда преобладает тета-ритм.
Почему мозг любит создавать тета-ритм?
Почему же мозг так любит создавать тета-ритм? Для ответа на этот вопрос нужны данные, полученные другой методикой изучения электрической активности мозга.
Опущенные в мозг микроэлектроды (стеклянные капилляры или изолированные на поверхности вольфрамовые проволочки) с кончиками всего в несколько микрон позволили прослушать работу отдельных нервных клеток — нейронов. Их «разговор» отличается от «шума» тех мозговых волн, чьи ритмы мы обсуждали выше. Импульс, или как его называют, спайк, длящийся одну миллисекунду, является тем словом, которое, с разной частотой повторяясь, составляет «речь» каждого нейрона.
Оказалось, что на любое раздражение, приходящее в мозг, многие нейроны, если они возбуждаются, отвечают по одной схеме: дают несколько спайков, замолкают и вновь начинают разряжаться (иногда даже чаще, чем в первый раз) через 140—250 миллисекунд. Приведенные цифры составляют как раз длительность одной волны тета-ритма. Можно предположить, что вовлечение новых нейронов в реакцию наиболее успешно происходит именно в то время, когда возникает наиболее сильный — второй период импульсации, и потому распространение возбуждения особенно эффективно при наличии тета-ритма.

Принципиальная схема машинного топоскопа (прибора многоканальной регистрации биопотенциалов) на 24 отведения и автоматизированного управляемого эксперимента на кролике. Сверху дан контур полушария мозга кролика, где кружками обозначены электроды. Спускаясь вниз, мы видим топоскоп, коммутатор, аналого-цифровой преобразователь и электронно-цифровую вычислительную машину (ЭЦВМ), от которой влево отходит печатающее устройство (сюда выдаются числовые значения коэффициентов кросскорреляции между биопотенциалами различных точек мозговой коры), а вправо — устройство согласования с объектом (УСО). От УСО по стрелке можно проследовать наверх, где обозначен световой раздражитель и глаз животного. Слева от контура мозга нарисована лапа, с которой регистрируется электро-миограмма

Кролик с микроэлектродами для записи потенциалов отдельных нервных клеток (нейронов)

Установка для автоматизированного управляемого эксперимента на кролике
Следовательно, результаты, полученные с помощью микроэлектродной техники как в нашей лаборатории (здесь в первую очередь нужно отметить опыты И. Н. Кондратьевой), так и многими другими исследователями, свидетельствуют о том, что корковым нейронам «удобнее» разряжаться в тета-ритме. Эти данные характеризуют, так сказать, оптимальную длину шага марширующего на параде солдата. А где расположен тот оркестр, тот ритмоводитель, который придает четкость движению всех колонн?
До сих пор, говоря о пространственной синхронизации биопотенциалов, мы ограничивались корой больших полушарий мозга, так как именно с коры регистрировали эти биопотенциалы. Но кора является частью мозга, которая тесно связана с другими, подкорковыми отделами, и не исключена возможность, что регистрируемые с ее поверхности электрические волны являются отражением глубинных процессов.
Тета-ритм в коре и подкорковой структуре мозга
Когда речь заходит о тета-ритме, перед мысленным взором нейрофизиолога возникает подкорковая структура мозга, очертания которой напоминают морского конька.
Дело в том, что сильнее всего тета-ритм проявляется в гиппокампе. В тех участках коры, которые расположены над гиппокампом, он может появляться в результате простого физического затекания. Но этот ритм может продуцироваться и в самой коре.
Детальные исследования по этой проблеме были проведены в нашей лаборатории В. Д. Трушом и Т. А. Корольковой. Было выяснено, что синхронность между биопотенциалами коры и гип-покампа тем выше, чем ближе корковое отведение к гиппокампу. По мере удаления от гиппокампа (а он располагается под ограниченной областью коры) синхронность между ним и коркой резко уменьшается. Одно из возможных и наиболее простых объяснений такой зависимости заключается в том, что некоторая доля электрической активности, отводимой от коры, определяется электрическим полем гиппокампа. Но так ли это?
Когда определяли одновременно синхронность биопотенциалов в двух точках коры и в двух точках гиппокампа, то обнаружили, что при высокой синхронности и в биопотенциалах корковых точек в гиппокампальных отведениях синхронность была высокой только в 5,7% случаев.
Если же была велика синхронность в гиппокампальных пунктах, то в корковых значительная синхронность возникала тоже не часто — только в 14,3 % случаев.
Решающими оказались данные, полученные с помощью управляемого эксперимента. При высокой синхронности между биопотенциалами зрительной и двигательной областей коры движения на свет возникали чаще, чем при высокой синхронности между биопотенциалами коры и гиппокампа.
Следовательно, даже при переводе с «языка» простых движений кролика на «язык» биопотенциалов головного мозга надо различать разные мозговые структуры, в которых пространственная синхронизация биопотенциалов может возникать по разным причинам. И даже когда мы точно знаем источник биопотенциалов, то еще остается открытым вопрос: «говорит» ли этот источник сам или только отражает чужую «речь»?
Вероятно, нужно признать, что функционально значимое повышение синхронности корковых биопотенциалов обусловлено прежде всего активным состоянием самой коры, хотя физическое затекание тета-ритма на кору из гиппокампа тоже не исключается.
Как расчленить эти источники? В опытах на кроликах было предпринято радикальное решение вопроса, а именно: небольшой участок коры (в дальнейшем называемой изолированной полоской) хирургическим путем лишали всех нервных связей с остальным мозгом.
Однако топография полоски не менялась, она оставалась в том же месте внутри черепа и была связана с остальным организмом через кровеносные сосуды-оболочки мозга. На нейронально изолированной полоске коры, где собственная электрическая активность из-за нарушения всех нервных связей с остальным мозгом несколько заглушена, наблюдали физическое (а не по нервным путям) затекание тета-ритма от гиппокампа. Однако и в самой полоске коры регистрировали собственный тета-ритм.
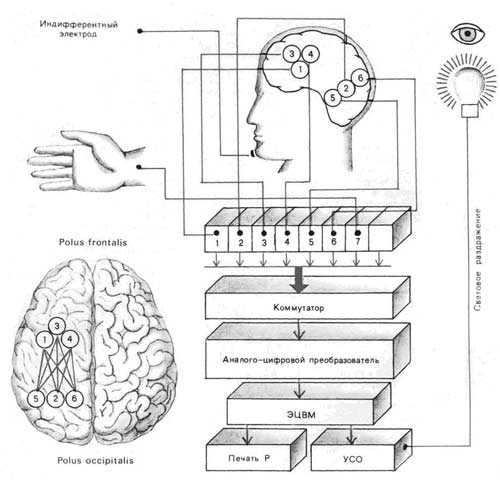
Принципиальная схема управляемого эксперимента на человеке и расположение электродов на поверхности черепа
Более детально пространственную синхронизацию на этом препарате и на более крупных изолированных мозговых структурах изучала наша сотрудница Л. В. Тишанинова. Синхронность биопотенциалов разных отделов нейронального изолированного полушария головного мозга кролика в сравнении с другим нетронутым полушарием возрастала, но была менее длительной. Кроме того, исчезали компактные зоны пространственной синхронности. Следовательно, одни только внутри-корковые механизмы могут обеспечивать определенный уровень пространственной синхронности, но для нормальной организации процессов необходима сохранность связей коры с подкорковыми центрами.
Если рассмотренные нами пространственно-временные взаимоотношения биопотенциалов различных участков головного мозга могут более полно описать его деятельность, то важно было проверить, как они изменяются при образовании условных рефлексов.
Было отмечено, что пространственная синхронизация биопотенциалов усиливается при выработке электрооборонительного условного рефлекса у кролика: в ответ на световой или звуковой раздражитель животное должно было отдернуть лапу, перед тем как получить электрическое раздражение этой конечности. Ко времени появления условного рефлекса синхронизация широко охватывает кору, обеспечивая переход возбуждения с корковой зоны представительства условного раздражителя к двигательному центру. Важно, что усиление синхронизации биопотенциалов происходит в диапазоне тета-ритма, т. е. в диапазоне частот от 4 до 7 гц. Причем процессы в разных точках мозга реализуются не только с одинаковой частотой, но совпадают и по форме каждой волны. Иными словами, укрепление условного рефлекса сопровождается повышением когерентности и синфазности колебаний биопотенциалов коры головного мозга в диапазоне тета-ритма. Даже в межсигнальные периоды (когда нет подачи условных раздражителей) в начале выработки временной связи отмечается усиление когерентности, а затем ее ослабление по мере укрепления условного рефлекса. Но и в этот период при осуществлении уcловнорефлекторного движения резко повышается когерентность в диапазоне тета-ритма.
Как при анализе простых движений, так и при анализе формирования временных связей необходимо было проверить взаимоотношения биопотенциалов мозга не только в одной плоскости (т. е. в коре), но и по глубине.
Важность глубинных (подкорковых) образований
Важность глубинных (подкорковых) образований в организации пространственной синхронизации биопотенциалов коры при образовании условных рефлексов была обнаружена в опытах нашей сотрудницы И. Н. Книпст. Она вживляла десятки проволочек-электродов в самые различные области головного мозга кролика (ретикулярная формация среднего мозга, гипоталамус, таламус, кора и др.). Животное с такими электродами может жить длительное время, не обнаруживая в своем поведении заметных отклонений от нормы. При сравнении степени синхронности биопотенциалов в разных участках мозга было обнаружено, что в период укрепления условного рефлекса усиливалась синхронность как в коре, так и между разными подкорковыми и, наконец, между подкоркой и корой.

Спектральная характеристика биопотенциалов мозга кролика, отводимых (сверху вниз) от гиппокампа, двигательной коры и зрительной коры. Каждая кривая — результат анализа четырехсекундной записи биопотенциалов мозга. На абциссе — частота в герцах, на ординате — выраженность каждой частоты в относительных единицах; размерность приведена в левом нижнем углу. Первая колонка кривых характеризует фоновую запись, а последующие показывают частотную характеристику биопотенциалов при действии световых вспышек с частотой 2, 3, 5, 7 и 10 гц. Можно видеть, что по выраженности и по степени сходства биопотенциалов, отводимых от различных участков мозга, выделяются данные, полученные при засвете с частотой 5 гц
Следовательно, относительно независимые биоэлектрические процессы коры и подкорки в результате сочетания условного раздражителя с безусловным сдвигаются в сторону пространственной синхронизации этих процессов и формирования корково-подкорко-вых синхронно действующих систем. Интересно, что корково-подкорковые связи при этом были распределены неравномерно, и одни корковые области «проявляли симпатию» к определенным подкорковым структурам, а другие нет. На основании изучения этих подвижных соотношений можно сделать вывод, что при учете ведущего значения коры головного мозга в становлении условного рефлекса (особенно в начальной стадии) существенную роль играют задние отделы гипоталами-ческой области и ретикулярная формация среднего мозга. Позднее, когда условнорефлекторный навык полностью устанавливается, отмечается вовлечение гип-покампа при одновременном ослаблении активации ретикулярной формации.
Конечно, представленная картина не претендует на исчерпывающее описание отношений между корой и подкоркой при формировании временных связей, но она может служить основанием для дальнейших исследований в этой области.
Синхронность биопотенциалов при возникновении торможения
Все сказанное выше характеризует один из основных нервных процессов — процесс возбуждения. А как изменяется синхронность биопотенциалов при возникновении торможения?
В нашей лаборатории Л. А. Потуловой был исследован этот вопрос при введении кроликам в различных дозах снотворного вещества — амитала натрия. Спектральный анализ (анализ спектра частот) показал, что по мере углубления сна происходит снижение доминирующих частот иногда до дельта-диапазона (0,5—3,0 гц). Наступает снижение кросскорреляционных коэффициентов при сравнении биопотенциалов зрительной и двигательной областей коры, и особенно между корой и подкорковыми центрами. В основе этого явления лежат расхождение частоты биопотенциалов в разных областях и увеличение их фазового сдвига. Оба эти процесса ведут к функциональной изоляции корковых зон друг от друга и прежде всего к разобщению деятельности коры и подкорковых обра зований головного мозга.
Когда у кроликов вырабатывали дифференцировочное торможение (опыты Т. М. Ефремовой и В. Д. Труша), т. е. подкрепляли электрическим током действие одного раздражителя и не подкрепляли действие другого, то на неподкрепляемый (дифференци-ровочный) стимул возникало снижение согласованности (когерентности) биоэлектрических колебаний в диапазоне 4—7 гц. Напомним, что действие подкрепляемого (положительного) стимула вызывало, напротив, резкое повышение когерентности. При другом виде тормозного процесса — при угасательном торможении, возникающем в случае отмены подкрепления условного раздражителя (вследствие этого исчезает рефлекс), также наблюдали расхождение доминирующих частот в диапазоне тета-ритма, т. е. функциональное рассогласование соответствующих отделов головного мозга.
В основе всех трех исследуемых видов торможения (сонное, дифференцировочное, угасательное) лежит, видимо, сходный процесс снижения возможности распространения возбуждения в головном мозге. При этом наступает расхождение частот в диапазоне тета-ритма, а также падение когерентности и синфаз-ности медленных биоэлектрических колебаний. Таким образом, было показано, что суть явления заключается в своеобразном «разобщении» процессов, протекающих в различных корковых и, вероятно, подкорковых центрах.
Вновь возвращаясь к сравнению с лингвистикой, мы можем заключить, что на новом языке, отражающем пространственно-временную характеристику биопотенциалов головного мозга, можно достаточно успешно описать такие важные нейрофизиологические явления, как распространение возбуждения, формирование тормозного процесса или замыкание условных рефлексов.
Отсутствии строгой локализации нервных и психических процессов
Заканчивая, мы должны отметить, что важным итогом современных нейрофизиологических исследований является вывод об отсутствии строгой локализации многих нервных и психических процессов. Погоня за центрами памяти, сна, сознания и т. п. не увенчалась успехом.
Метод множественной регистрации биоэлектрической активности мозга позволяет по-новому оценивать нервную деятельность целостного мозга (Один из примеров — использование этого метода для изучения психической деятельности человека. См. статью Н. П. Бехтеревой и П. В. Бундзена «Нейрофизиология психической деятельности человека», опубликованную в ежегоднике «Наука и человечество. 1974». — Ред.).
Этот метод как бы расширяет горизонт исследователя и позволяет уже на стадии эксперимента непосредственно осуществлять столь необходимое в наше время обобщение данных, характеризующих местные нервные процессы. Перспективность этого пути уже ясна, и дальнейшие успехи будут связаны с увеличением числа отводимых пунктов и совершенствованием ЭВМ. Здесь прогресс может во многом обеспечиваться техническими средствами.
Кроме этого, существенное значение имеет совершенствование математических методов обработки получаемых обширных экспериментальных данных. Выделение наиболее информативных компонентов из записи биоэлектрической активности мозга является важной задачей, решаемой в совместной работе математиками и физиологами.
Наконец, новая форма исследования — управляемый эксперимент — открывает большие перспективы для более глубокого исследования деятельности мозга.
Эта форма эксперимента, основанная на глубоком знании физиологических фактов и умелом использовании возможностей современной вычислительной техники, позволит раскрыть многие тайны мозга.
Михаил Николаевич Ливанов
нейрофизиолог, акдемик,
Юрий Андреевич Холодов
нейрофизиолог, доктор биологических наук.
Пространственная синхронизация мозговых ритмов // Наука и человечество. 1975. Сборник. - М., Знание, 1974. С. 13–25.