
Мембранные процессы
Биологические мембраны, находящиеся на границе клетки и внеклеточного пространства, а также на границе мембранных органелл клетки (митохондрий, эндоплазматической сети, комплекса Гольджи [1], лизосом, пероксисом, ядра, мембранных пузырьков) и цитозоля существенно важны для функционирования как клетки в целом, так и её органелл. Клеточные мембраны имеют принципиально сходную молекулярную организацию. В этой главе биологические мембраны рассмотрены преимущественно на примере плазматической мембраны (плазмолеммы), отграничивающей клетку от внеклеточной среды.
Плазматическая мембрана
Любая биологическая мембрана (рис. 2–1) состоит из фосфолипидов (~50%) и белков (до 40%). В меньших количествах в состав мембраны входят другие липиды, холестерол и углеводы.

Рис. 2–1. Биологическая мембрана состоит из двойного слоя фосфолипидов, гидрофильные части которых (головки) направлены к поверхности мембраны, а гидрофобные части (хвосты, стабилизирующие мембрану в виде бислоя) внутрь мембраны. И — интегральные белки погружены в мембрану. Т — трансмембранные белки пронизывают всю толщу мембраны. П — периферические белки расположены либо на наружной, либо на внутренней поверхности мембраны.
Фосфолипиды. Молекула фосфолипида состоит из полярной (гидрофильной) части (головка) и аполярного (гидрофобного) двойного углеводородного хвоста. В водной фазе молекулы фосфолипидов автоматически агрегируют хвост к хвосту, формируя каркас биологической мембраны (рис. 2–1 и 2–2) в виде двойного слоя (бислой). Таким образом, в мембране хвосты фосфолипидов (жирные кислоты) направлены внутрь бислоя, а содержащие фосфатные группировки головки обращены кнаружи.
Арахидоновая кислота. Из мембранных фосфолипидов освобождается арахидоновая кислота — предшественник Пг, тромбоксанов, лейкотриенов и ряда других биологически активных веществ с множеством функций (медиаторы воспаления, вазоактивные факторы, вторые посредники и др.).
Липосомы — искусственно приготовленные из фосфолипидов мембранные пузырьки диаметром от 25 нм до 1 мкм. Липосомы используют как модели биологических мембран, а также для введения внутрь клетки различных веществ (например, генов, ЛС); последнее обстоятельство основано на том, что мембранные структуры (в т.ч. и липосомы) легко сливаются (за счёт фосфолипидного бислоя).
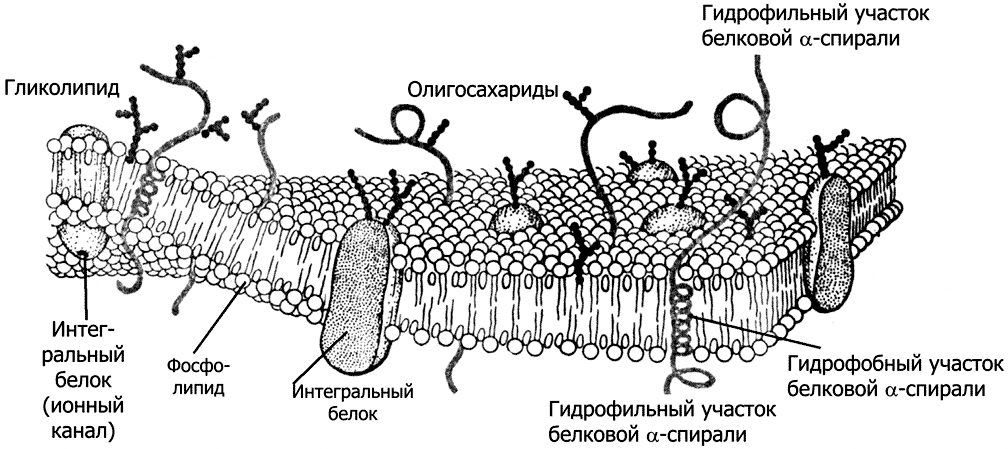
Рис. 2–2. Плазматическая мембрана.
Белки биологических мембран подразделяют на интегральные (в том числе трансмембранные) и периферические (рис. 2–1 и 2–2).
Интегральные мембранные белки (глобулярные) встроены в липидный бислой. Их гидрофильные аминокислоты взаимодействуют с фосфатными группами фосфолипидов, а гидрофобные аминокислоты — с цепями жирных кислот. К интегральным мембранным белкам относятся белки адгезии, некоторые рецепторные белки (мембранные рецепторы).
Трансмембранный белок — молекула белка, проходящая через всю толщу мембраны и выступающая из неё как на наружной, так и на внутренней поверхности. К трансмембранным белкам относятся поры, ионные каналы, переносчики, насосы, некоторые рецепторные белки.
Поры и каналы — трансмембранные пути, по которым между цитозолем и межклеточным пространством (и в обратном направлении) перемещаются вода, ионы и молекулы метаболитов.
Переносчики осуществляют трансмембранное перемещение конкретных молекул (в том числе в сочетании с переносом ионов или молекул другого типа).
Насосы перемещают ионы против их концентрационного и энергетического градиентов (электрохимический градиент) при помощи энергии, освобождаемой при гидролизе АТФ.
Периферические мембранные белки (фибриллярные и глобулярные) находятся на одной из поверхностей клеточной мембраны (наружной или внутренней) и нековалентно связаны с интегральными мембранными белками.
Примеры периферических мембранных белков, связанных с наружной поверхностью мембраны — рецепторные белки и белки адгезии.
Примеры периферических мембранных белков, связанных с внутренней поверхностью мембраны, — белки цитоскелета, белки системы вторых посредников, ферменты и другие белки.
Латеральная подвижность. Интегральные белки могут перераспределяться в мембране в результате взаимодействия с периферическими белками, элементами цитоскелета, молекулами в мембране соседней клетки и компонентами внеклеточного матрикса.
Углеводы (преимущественно олигосахариды) входят в состав гликопротеинов и гликолипидов мембраны, составляя 2–10% её массы (рис. 2–2). С углеводами клеточной поверхности взаимодействуют лектины. Цепи олигосахаридов выступают на наружной поверхности мембран клетки и формируют поверхностную оболочку — гликокаликс.
Гликокаликс имеет толщину около 50 нм и состоит из олигосахаридов, ковалентно связанных с гликопротеинами и гликолипидами плазмолеммы. Функции гликокаликса: межклеточное узнавание, межклеточные взаимодействия, пристеночное пищеварение (гликокаликс, покрывающий микроворсинки каёмчатых клеток эпителия кишечника, содержит пептидазы и гликозидазы, завершающие расщепление белков и углеводов).
Проницаемость мембраны
Мембранный бислой разделяет две водные фазы. Так, плазматическая мембрана отделяет межклеточную (интерстициальную) жидкость от цитозоля, а мембраны лизосом, пероксисом, митохондрий и других мембранных внутриклеточных органелл их содержимое от цитозоля.[2] Биологическая мембрана — полупроницаемый барьер.
Полупроницаемая мембрана. Биологическую мембрану определяют как полупроницаемую, т.е. барьер, не проницаемый для воды, но проницаемый для растворённых в ней веществ (ионы и молекулы).
Полупроницаемые тканевые структуры. К полупроницаемым тканевым структурам относят также стенку кровеносных капилляров и различные барьеры (например, фильтрационный барьер почечных телец, аэрогематический барьер респираторного отдела лёгкого, гематоэнцефалический барьер и многие другие, хотя в состав таких барьеров — помимо биологических мембран (плазмолемма) — входят и немембранные компоненты. Проницаемость таких тканевых структур рассмотрена в разделе «Трансклеточная проницаемость» главы 4.
Физико-химические параметры межклеточной жидкости и цитозоля существенно различны (см. табл. 2-1), также различны параметры каждого мембранного внутриклеточного органоида и цитозоля. Наружная и внутренняя поверхности биологической мембраны полярны и гидрофильны, но неполярная сердцевина мембраны гидрофобна. Поэтому неполярные вещества могут проникать через липидный бислой. В то же время именно гидрофобный характер сердцевины биологической мембраны определяет принципиальную невозможность непосредственного проникновения через мембрану полярных веществ.
Неполярные вещества (например, водонерастворимые холестерол и его производные) свободно проникают через биологические мембраны. В частности, именно по этой причине рецепторы стероидных гормонов расположены внутри клетки.
Полярные вещества (например, ионы Na+, K+ C1- , Са2+; различные небольшие, но полярные метаболиты, а также сахара, нукле-отиды, макромолекулы белка и нуклеиновых кислот) сами по себе не проникают через биологические мембраны. Именно поэтому рецепторы полярных молекул (например, пептидных гормонов) встроены в плазматическую мембрану, а передачу гормонального сигнала к другим клеточным компартментам осуществляют вторые посредники.
Избирательная проницаемость — проницаемость биологической мембраны по отношению к конкретным химическим веществам) –– важна для поддержания клеточного гомеостаза. оптимального содержания в клетке ионов, воды, метаболитов и макромолекул. Перемещение конкретных веществ через биологическую мембрану называют трансмембранным транспортом (чрезмембранный транспорт).
Орлов Р.С., Ноздрачёв А.Д. Нормальная физиология. – М., 2009. – 688 с.
Глава 2. Мембранные процессы. С. 15-18.
[1] Гольджи Камилло (Golgi C.), итальянский гистолог (1843–1926). Разработал метод окрашивания ткани, основанный на импрегнации соединениями металлов. Метод был развит Рамон-и-Кахалем и его учениками, что позволило описать строение практически всех отделов нервной системы. За разработку этого метода Гольджи в 1906 г. был удостоен Нобелевской премии по медицине вместе с Сантъяго Рамон-и-Кахалем.
[2] Цитозоль (англ. cytosol) или внутриклеточная жидкость, матрикс цитоплазмы, гиалоплазма — жидкость, находящаяся внутри клеток. У эукариот матрикс цитоплазмы отделен клеточными мембранами от содержимого органоидов, например, матрикса митохондрий. Содержимое клетки за исключением плазматической мембраны и ядра называют цитоплазмой. [Прим. reabilitaciya.org]