
Транспортёры специфичны: каждый конкретный переносчик переносит через липидный бислой, как правило и по преимуществу, одно вещество. Различают однонаправленный (унипорт), сочетанный (симпорт) и разнонаправленный (антипорт) транспорт (рис. 2–6).

Рис. 2–6 (CD-ROM). Модель вариантов трансмембранного переноса разных молекул [9].
Унипорт — однонаправленный транспорт одного вещества (рис. 2–6).
Симпорт (котранспорт). Некоторые переносчики осуществляют однонаправленный сочетанный транспорт одного вещества (рис. 2–6).
Антипорт (обменник). Эти переносчики осуществляют разнонаправленный транспорт двух разных веществ (встречный транспорт, рис. 2–6).
Вторичный активный транспорт. Переносчики, реализующие как сочетанный (симпорт), так и разнонаправленный (антипорт) трансмембранный перенос с точки зрения энергетических затрат функционируют так, что энергия, накапливающаяся при переносе одного вещества (обычно Na+), затрачивается на транспорт другого вещества. Такой тип трансмембранного переноса называют вторичным активным транспортом (см. ниже).
Кинетика переноса (рис. 2–7, см. также рис. 2–3Б), как и скорость ферментной реакции, описывается уравнением Михаэлиса–Ментена, но вместо скорости ферментной реакции подставлено значение J (величина трансмембранного переноса):
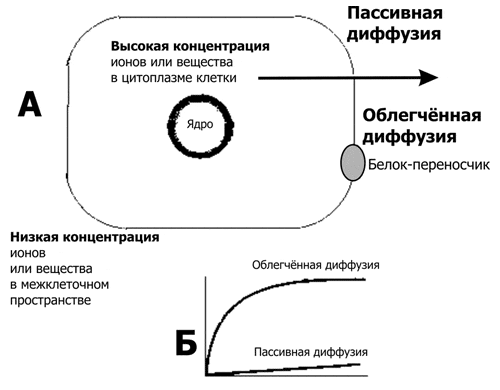{kind=link}
Уравнение 2–4
JX = ([X]Jмакс) ? (Km + X)
где Х — транспортируемое вещество,
JX — величина транспорта вещества Х,
Jмакс — максимальная величина транспорта,
Km — константа Михаэлиса (численно равна концентрации вещества Х, когда JX составляет половину Jмакс).

Рис. 2–7 (CD-ROM). Модель взаимодействия переносчика и транспортируемой молекулы Х [2]. Белки переносчика формируют канал, на внутренней поверхности которого имеется участок связывания молекулы Х (выемка). Диффузия молекул Х через мембрану в принципе может происходить в обеих направлениях, направление определяется концентрационным или электрохимическим трансмембранным градиентом для вещества Х. На рисунке показаны этапы (1–6) переноса молекулы Х слева (межклеточная среда) направо (цитозоль). Канал переносчика может находиться в открытом наружу (1, 2), внутрь клетки (4, 5) или закрытом (3, 6) состоянии.
Примеры трансмембранных переносчиков
Глюкоза
Унипорты глюкозы GLUT (от англ. GLUcose Transporter) — интегральные гликопротеины. Клетки, имеющие значительную потребность в глюкозе (в первую очередь нейроны ЦНС), содержат переносчик глюкозы. Инсулин увеличивает захват глюкозы клетками, вызывая быстрое перемещение переносчиков из цитоплазмы клетки в плазмолемму. В кардиомиоцитах экспрессию гена GLUT4 усиливают йодсодержащие гормоны щитовидной железы. GLUT5 щёточной каёмки энтероцитов тонкого кишечника обеспечивает всасывание глюкозы (также фруктозы), а GLUT2 базолатеральной части энтероцитов осуществляет выход этих сахаров из энтероцитов во внутреннюю среду организма. Точечная мутация гена GLUT2 (замена валина на изолейцин в позиции 197) — одна из причин развития сахарного диабета типа II.
Симпорт. Сочетанный транспорт глюкозы и Na+ в тонком кишечнике и канальцах почки обеспечивают мембранные гликопротеины, кодируемые генами SGLT. Это главный механизм почечной реабсорбции глюкозы, происходящей в начальном отделе проксимальных извитых канальцев нефрона. Известно несколько мутаций гена SGLT2, приводящих к потере глюкозы (глюкозурия) в почках (до 60 г/сут).
Аминокислоты
Всасывание аминокислот в кишечнике, их реабсорбция в канальцах нефрона, а также поглощение аминокислот–нейромедиаторов нейронами и глиоцитами мозга реализуются при помощи переносчиков, кодируемых генами SLC1 и SLC3. Идентифицированы десятки переносчиков, специфичных по отношению к a-, двухосновным и нейтральным аминокислотам.
Цистинурия. Мутации гена SLC3A1 приводят в почечных канальцах к избыточной секреции цистина (при некоторых формах также лизина, аргинина и орнитина) и к образованию мочевых камней.
Глутамат и аспартат (аминокислоты–нейромедиаторы), накапливаясь в межклеточном пространстве ЦНС, могут оказывать цитотоксический эффект. Кодируемый геном SLC1A2 белок–переносчик транспортирует эти аминокислоты в цитоплазму нейронов и глиоцитов.
Анионообменники
Анионообменники — регуляторы внутриклеточного pH, осуществляют обмен Cl– на бикарбонат.
«Полоса 3 мембраны эритроцитов» — многофункциональный транспортный белок глюкозы, анионов и воды.
Респираторный обмен CO2. Этот анионообменник обеспечивает эффективный обмен Cl– на HCO3–: поступление бикарбоната в эритроциты в обмен на Cl– в капиллярах разных органов, обратный процесс происходит в капиллярах лёгкого.
Аг старения клеток. При деградации этого белка образуется т.н. Аг старения клеток — метка, распознаваемая иммунной системой. Эритроциты, несущие эту метку (повреждённые, заканчивающие жизненный цикл, инфицированные плазмодиями малярии), распознаются макрофагами селезёнки и печени и фагоцитируются ими.
Фосфаты. Реабсорбция фосфатов происходит в проксимальных извитых канальцах нефрона при помощи переносчиков фосфатов.
Обмен Na+ на H+. Гены SLC9A кодируют белки плазмолеммы NHE (от Na, H, Exchanger [обменник]), осуществляющие обмен внеклеточного Na+ на внутриклеточный H+. Мембранные белки NHE — регуляторы внутриклеточного pH. Диуретик амилорид ингибирует NHE1, NHE2 и NHE4 и не оказывает эффекта на NHE3 и NHE5.
Разные переносчики
Креатин. Кардиомиоциты и скелетные МВ не синтезируют креатина, необходимого для энергетического обеспечения их функции. Ингибирование переносчика креатина ведёт к развитию мышечной слабости. Дефекты кодирующего переносчик гена SLC6A8 — причина одной из форм кардиомиопатии.
Фолаты. Этот переносчик обнаружен в плаценте и печени.
Лактат и пируват транспортируются в обоих направлениях через плазмолемму эритроцитов, гепатоцитов, эпителия кишечника и почки, клеток мышечных тканей.
Норадреналин. [1] Выделившиеся в синаптическую щель нейромедиаторы частично транспортируются обратно в пресинаптические терминали. Трициклические антидепрессанты (например, дезипрамин, имипрамин), а также кокаин и амфетамины взаимодействуют с синаптическими транспортными системами биогенных аминов. Мутации генов, кодирующих переносчики норадреналина и серотонина [2],могут привести к развитию психиатрической патологии (например, маниакально–депрессивного психоза).
Орлов Р.С., Ноздрачёв А.Д. Нормальная физиология : Учебник. – М., 2009. – 688 с.
Глава 2. Мембранные процессы. С. 20, CD-ROM.
[1] Норадреналин — деметилированный предшественник адреналина — 2-амино-1-(3,4-дигидроксифенил) этанол. Секретируется из большинства постганглионарных симпатических волокон и является н. между многими нейронами ЦНС (например, гипоталамус, голубоватое место). Образуется из дофамина путём гидролиза при помощи дофамин-b-гидроксилазы. Норадреналин хранится в синаптических пузырьках, после высвобождения взаимодействует с адренорецепторами, реакция прекращается в результате захвата норадреналина пресинаптической частью. Уровень норадреналина определяется активностью тирозин гидроксилазы и моноаминооксидазы. Моноаминооксидаза и катехол-О-метилтрансфераза переводят норадреналин в неактивные метаболиты (норметанефрин, 3-метокси-4-гидрокси-фенилэтиленгликоль, 3-метокси-4-гидроксиминдальная кислота). Норадреналин — мощный вазоконстриктор, эффект происходит при взаимодействии норадреналина с ГМК стенки кровеносных сосудов. Транспортёры. Захват норадреналина в межклеточном пространстве (синаптической щели) осуществляют специфические Na+- и Cl–-транспортирующие белки (например, норадреналин-транспортирующий белок 1. Эти белки — мишени трициклических антидепрессантов (например, дезипрамин и имипрамин). Система захвата биогенных аминов — точка приложения антидепрессантов и таких препаратов, как кокаин и амфетамины. Дефекты транспортёров норадреналина и серотонина — кандидаты на роль первопричины при психиатрических расстройствах, таких как маниакально-депрессивные состояния/ [2] Серотонин (5-гидрокситриптамин) образуется в тромбоцитах и других клетках из 5?окситриптофана в результате его декарбоксилирования неспецифической декарбоксилазой ароматических L?аминокислот. Сосудосуживающее вещество, ингибитор секреции желудочного сока, стимулятор сокращения ГМК; присутствует в относительно высоких концентрациях в некоторых областях ЦНС (серотонин — нейромедиатор многих центральных нейронов, например ядра шва), многих периферических тканях и клетках, а также в опухолях. Расщепляется серотонин моноаминооксидазой с образованием 5?гидроксииндолуксусной кислоты. Депрессия характеризуется снижением количества норадреналина и серотонина и увеличением экспрессии их рецепторов. Антидепрессанты уменьшают число этих рецепторов. Маниакальный синдром. При этом состоянии увеличивается уровень норадреналина на фоне снижения количества серотонина и адренорецепторов. Литий снижает секрецию норадреналина, образование вторых посредников и увеличивает экспрессию адренорецепторов. Аутизм. Гиперсеротонинемия, но в 30–50% случаев без явных нарушений обмена серотонина в мозге.