
Тета-ритм в коре и подкорковой структуре мозга
Когда речь заходит о тета-ритме, перед мысленным взором нейрофизиолога возникает подкорковая структура мозга, очертания которой напоминают морского конька.
Дело в том, что сильнее всего тета-ритм проявляется в гиппокампе. В тех участках коры, которые расположены над гиппокампом, он может появляться в результате простого физического затекания. Но этот ритм может продуцироваться и в самой коре.
Детальные исследования по этой проблеме были проведены в нашей лаборатории В. Д. Трушом и Т. А. Корольковой. Было выяснено, что синхронность между биопотенциалами коры и гип-покампа тем выше, чем ближе корковое отведение к гиппокампу. По мере удаления от гиппокампа (а он располагается под ограниченной областью коры) синхронность между ним и коркой резко уменьшается. Одно из возможных и наиболее простых объяснений такой зависимости заключается в том, что некоторая доля электрической активности, отводимой от коры, определяется электрическим полем гиппокампа. Но так ли это?
Когда определяли одновременно синхронность биопотенциалов в двух точках коры и в двух точках гиппокампа, то обнаружили, что при высокой синхронности и в биопотенциалах корковых точек в гиппокампальных отведениях синхронность была высокой только в 5,7% случаев.
Если же была велика синхронность в гиппокампальных пунктах, то в корковых значительная синхронность возникала тоже не часто — только в 14,3 % случаев.
Решающими оказались данные, полученные с помощью управляемого эксперимента. При высокой синхронности между биопотенциалами зрительной и двигательной областей коры движения на свет возникали чаще, чем при высокой синхронности между биопотенциалами коры и гиппокампа.
Следовательно, даже при переводе с «языка» простых движений кролика на «язык» биопотенциалов головного мозга надо различать разные мозговые структуры, в которых пространственная синхронизация биопотенциалов может возникать по разным причинам. И даже когда мы точно знаем источник биопотенциалов, то еще остается открытым вопрос: «говорит» ли этот источник сам или только отражает чужую «речь»?
Вероятно, нужно признать, что функционально значимое повышение синхронности корковых биопотенциалов обусловлено прежде всего активным состоянием самой коры, хотя физическое затекание тета-ритма на кору из гиппокампа тоже не исключается.
Как расчленить эти источники? В опытах на кроликах было предпринято радикальное решение вопроса, а именно: небольшой участок коры (в дальнейшем называемой изолированной полоской) хирургическим путем лишали всех нервных связей с остальным мозгом.
Однако топография полоски не менялась, она оставалась в том же месте внутри черепа и была связана с остальным организмом через кровеносные сосуды-оболочки мозга. На нейронально изолированной полоске коры, где собственная электрическая активность из-за нарушения всех нервных связей с остальным мозгом несколько заглушена, наблюдали физическое (а не по нервным путям) затекание тета-ритма от гиппокампа. Однако и в самой полоске коры регистрировали собственный тета-ритм.
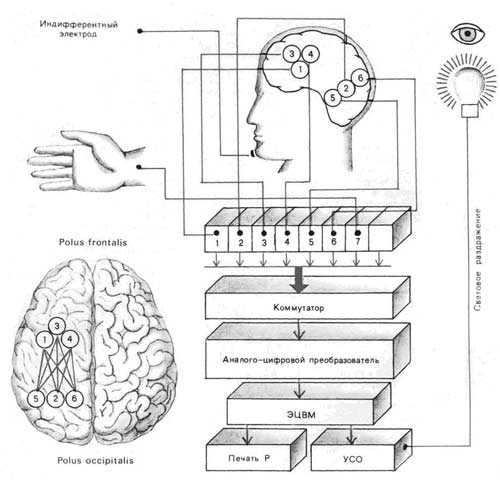
Принципиальная схема управляемого эксперимента на человеке и расположение электродов на поверхности черепа
Более детально пространственную синхронизацию на этом препарате и на более крупных изолированных мозговых структурах изучала наша сотрудница Л. В. Тишанинова. Синхронность биопотенциалов разных отделов нейронального изолированного полушария головного мозга кролика в сравнении с другим нетронутым полушарием возрастала, но была менее длительной. Кроме того, исчезали компактные зоны пространственной синхронности. Следовательно, одни только внутри-корковые механизмы могут обеспечивать определенный уровень пространственной синхронности, но для нормальной организации процессов необходима сохранность связей коры с подкорковыми центрами.
Если рассмотренные нами пространственно-временные взаимоотношения биопотенциалов различных участков головного мозга могут более полно описать его деятельность, то важно было проверить, как они изменяются при образовании условных рефлексов.
Было отмечено, что пространственная синхронизация биопотенциалов усиливается при выработке электрооборонительного условного рефлекса у кролика: в ответ на световой или звуковой раздражитель животное должно было отдернуть лапу, перед тем как получить электрическое раздражение этой конечности. Ко времени появления условного рефлекса синхронизация широко охватывает кору, обеспечивая переход возбуждения с корковой зоны представительства условного раздражителя к двигательному центру. Важно, что усиление синхронизации биопотенциалов происходит в диапазоне тета-ритма, т. е. в диапазоне частот от 4 до 7 гц. Причем процессы в разных точках мозга реализуются не только с одинаковой частотой, но совпадают и по форме каждой волны. Иными словами, укрепление условного рефлекса сопровождается повышением когерентности и синфазности колебаний биопотенциалов коры головного мозга в диапазоне тета-ритма. Даже в межсигнальные периоды (когда нет подачи условных раздражителей) в начале выработки временной связи отмечается усиление когерентности, а затем ее ослабление по мере укрепления условного рефлекса. Но и в этот период при осуществлении уcловнорефлекторного движения резко повышается когерентность в диапазоне тета-ритма.
Как при анализе простых движений, так и при анализе формирования временных связей необходимо было проверить взаимоотношения биопотенциалов мозга не только в одной плоскости (т. е. в коре), но и по глубине.